Purdue University College of Agriculture

Featured Stories

2016 was the first time that Jalyn Gearries, a Natural Resources and Environmental Science (NRES) alumna and Indiana native, ventured west of the Mississippi and experienced the Grand Canyon. This was the second stop on a three-week national park tour with her family, and it was that one stop that changed everything for her.

The Boilermaker Butcher Block’s selections will now include farm fresh eggs laid by Purdue chickens who are fed orange corn.
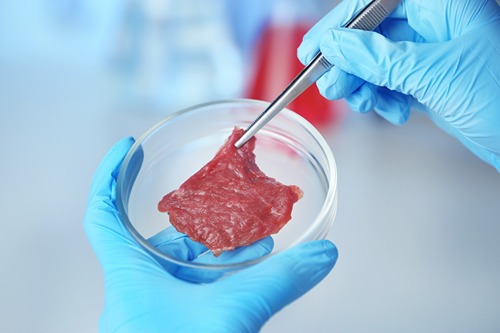
Many consumers view conventional meats as both tastier and healthier than laboratory-grown alternatives, according to the March Consumer Food Insights Report.
Prospective Students

What's Trending

PURDUE'S NEXT MOVES
In April 2021 Purdue launched five new distinct strategic initiatives designed to advance the University’s competitive advantage in its continuing quest for leadership among the world’s top research and teaching institutions.
Learn More
Purdue Extension
Purdue Extension is Indiana's educational partner for life. Extension delivers practical, research-based information that enhances lives and livelihoods.
Learn More